
Chapitre 5 Généralités sur les tumeurs Cellule et tissu cancéreux
Auteurs : Marius Ilié et Jean-François Emile
Plan du chapitre
• Définition d'une tumeur
• Bases moléculaires du cancer
• Modifications fonctionnelles et morphologiques
• Stroma tumoral
• Cancer et angiogenèse
• Immunité anti-tumorale
• Composition cellulaire d'une tumeur
• Tumeurs bénignes et malignes
• Nomenclature des tumeurs
• Stade et grade des tumeurs
• Méthodes diagnostiques des tumeurs
• Connaître la définition, la composition et la classification des tumeurs.
• Décrire les bases moléculaires de la cancérogenèse et connaître quelques exemples dans chacune des trois grandes familles de gènes (oncogènes, gènes suppresseurs et gènes de l'homéostasie génétique).
• Connaître les grands mécanismes de régulation de l'expression ou de la fonction de ces gènes.
• Connaître les principaux facteurs de risque génétiques et environnementaux des cancers.
• Connaître des exemples de dérégulation du cycle cellulaire et de l'apoptose dans les cancers.
• Décrire les caractéristiques biologiques et morphologiques d'une cellule cancéreuse. Décrire les cellules constituant généralement le stroma des tumeurs.
• Connaître les principales caractéristiques de la vascularisation des tumeurs
• Connaître les grands mécanismes de réponse immune antitumorale.
• Connaître les techniques histologiques et moléculaires utiles au diagnostic des tumeurs.
• Maîtriser les stratégies permettant de réaliser le diagnostic d'une tumeur.
La classification des tumeurs est fondée sur leur organe ou tissu d'origine, leur type histologique, et leur degré de malignité. Le diagnostic est fondé sur l'histologie, mais fait de plus en plus souvent appel à des techniques complémentaires telles que l'immunohistochimie, la cytogénétique et la pathologie moléculaire.
Définition d'une tumeur
La définition d'une tumeur repose sur la notion d'homéostasie tissulaire. Le développement et la croissance d'un tissu ou d'un organe sont conditionnés par des processus complexes permettant la régulation des différentes étapes de la vie d'une cellule : la prolifération, la différenciation, la sénescence et la mort cellulaire programmée. Tous ces phénomènes sont aussi mis en jeu lors du renouvellement des cellules dont la durée de vie est limitée. Au sein d'un tissu normal chez l'adulte, l'équilibre entre ces processus est à l'origine de l'homéostasie tissulaire. Les anomalies de cette homéostasie, par augmentation de la prolifération et/ou diminution de la mort cellulaire, sont à l'origine de l'accumulation des cellules aboutissant à la formation d'une tumeur macroscopiquement visible.
Il existe des tumeurs bénignes et des tumeurs malignes (cancers).
Le terme de tumeur (synonyme : néoplasme) désigne actuellement une prolifération cellulaire excessive aboutissant à une masse tissulaire ressemblant plus ou moins au tissu normal homologue (adulte ou embryonnaire), ayant tendance à persister et à croître, témoignant de son autonomie biologique.
Ces cellules tumorales dérivent dans la grande majorité des cas d'une seule cellule (monoclonales). Les particularités des cellules tumorales sont liées à l'accumulation d'altérations de leur génome (génotype). Ces altérations sont le plus souvent acquises au cours de la genèse tumorale, mais certaines peuvent être d'origine héréditaire (prédispositions familiales).
Les cellules tumorales peuvent acquérir de nouvelles altérations génétiques ou épigénétiques, et générer des sous-clones n'ayant conservé qu'une partie des caractéristiques morphologiques et fonctionnelles des cellules originelles et acquis de nouvelles (variabilité du phénotype des sous-clones).
Ces modifications peuvent s'inscrire dans le noyau, le cytoplasme ou la membrane des cellules tumorales, ainsi que dans leur agencement histologique ou leur stroma.
Caractères d'une tumeur
Prolifération cellulaire excessive
La prolifération est liée à la multiplication des descendants d'une ou plusieurs cellules anormales. C'est la notion de clonalité. Un clone est un ensemble de cellules dérivées d'une seule cellule initiale. Une tumeur est dite poly- oligo- ou monoclonale, selon qu'elle se développe à partir de plusieurs, de quelques ou d'une seule cellule.
Masse tissulaire ressemblant plus ou moins à un tissu normal
Les caractères cytologiques et architecturaux de ce nouveau tissu réalisent un aspect plus ou moins proche de celui du tissu normal homologue adulte ou embryonnaire. Cette ressemblance définit une notion fondamentale : la différenciation tumorale. Plus la fonction et la structure tumorales se rapprochent de la fonction et de la structure du tissu normal, plus la tumeur est dite différenciée (figure 05.01).
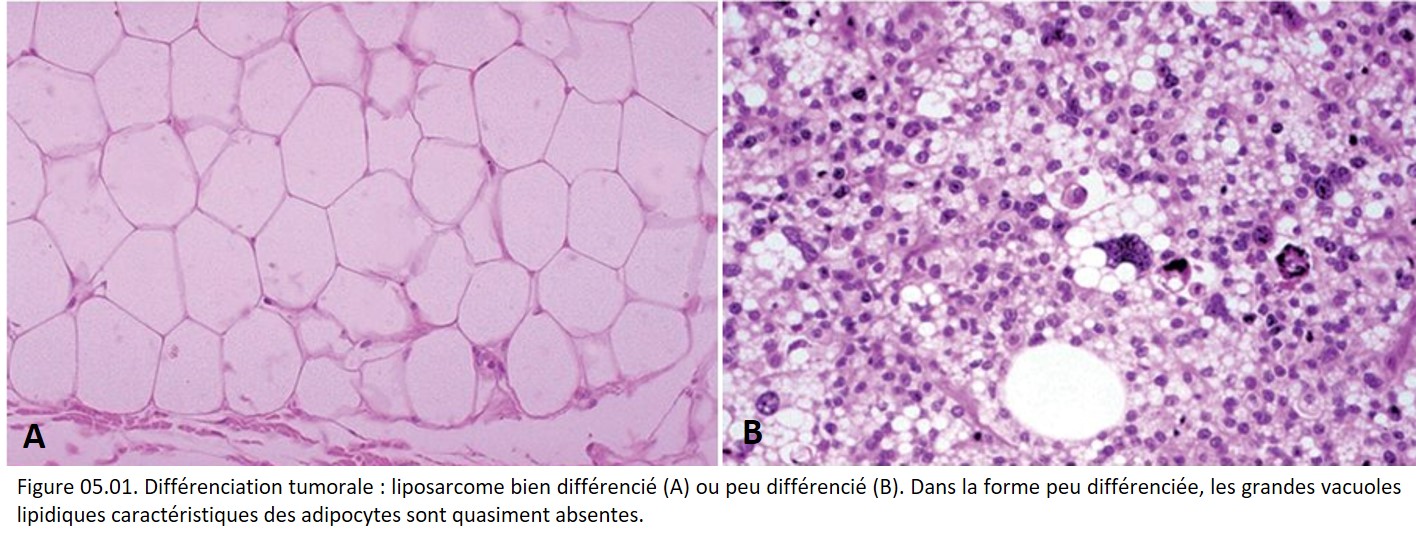{kind=link}
Tendance à persister et à croître
La prolifération tumorale se poursuit après la disparition du « stimulus » qui lui a donné naissance. La prolifération tumorale est biologiquement autonome.
Succession d'événements génétiques ou épigénétiques
Ces anomalies génétiques s'accumulent généralement en plusieurs années. Au cours de ce processus en plusieurs étapes, le génome des cellules tumorales acquiert des allèles mutants de proto-oncogènes, de gènes suppresseurs de tumeur et de gènes contrôlant directement ou indirectement l'intégrité de l'ADN.
La conséquence de ces anomalies génétiques est l'acquisition de nouvelles propriétés, dont la capacité :
• de générer leurs propres signaux mitogènes ;
• de résister ou d'échapper aux signaux externes d'inhibition de la croissance ;
• de proliférer sans limite (immortalisation) ;
• d'infiltrer les tissus adjacents et de disséminer à distance ;
• de constituer une néo-vascularisation (angiogénèse).
Bases moléculaires du cancer
Un cancer est la conséquence d'altérations successives du génome des cellules tumorales, qui perturbent de façon permanente l'homéostasie tissulaire (figure 05.02).
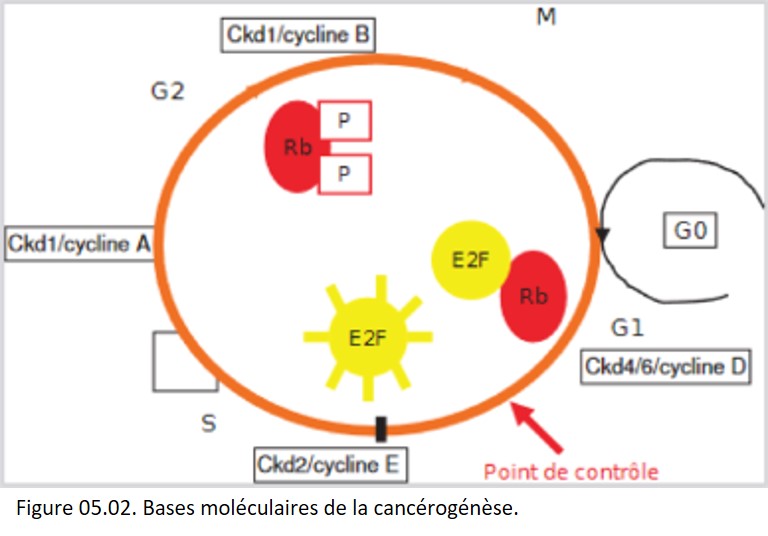{kind=link}
Dans la cellule cancéreuse, il y a rupture permanente de l'équilibre entre les signaux intracellulaires :
• activation de voies stimulatrices ;
• suppression de voies inhibitrices.
La coexistence de plusieurs événements est nécessaire à la transformation cancéreuse. Ce processus multi-étapes n'est pas une transition abrupte d'une croissance normale à une croissance tumorale, mais il se déroule progressivement sur plusieurs années, selon la courbe de croissance tumorale de Gompertz. Ces altérations sont le plus souvent acquises au cours de la genèse tumorale, mais certaines peuvent être d'origine héréditaire (prédispositions familiales) (ex. : certains cancers du sein ou du côlon). La survenue progressive de ces altérations sur plusieurs années constitue une opportunité pour le dépistage et le traitement des lésions précoces.
L'environnement extérieur joue également un rôle majeur. Ce rôle a été suspecté dés la fin du xviiie siècle par Percival Pott, chirurgien anglais, qui a décrit de fréquents cancers du scrotum chez de jeunes ramoneurs ayant commencé leur métier pendant l'enfance.
Différents agents de l'environnement conduisent au développement d'un cancer
• Agents initiateurs : ils induisent une lésion définitive de l'ADN (ex. : mutation, cassure). Souvent ces carcinogènes sont activés par des réactions métaboliques. Exemples (tableau 05.01) :
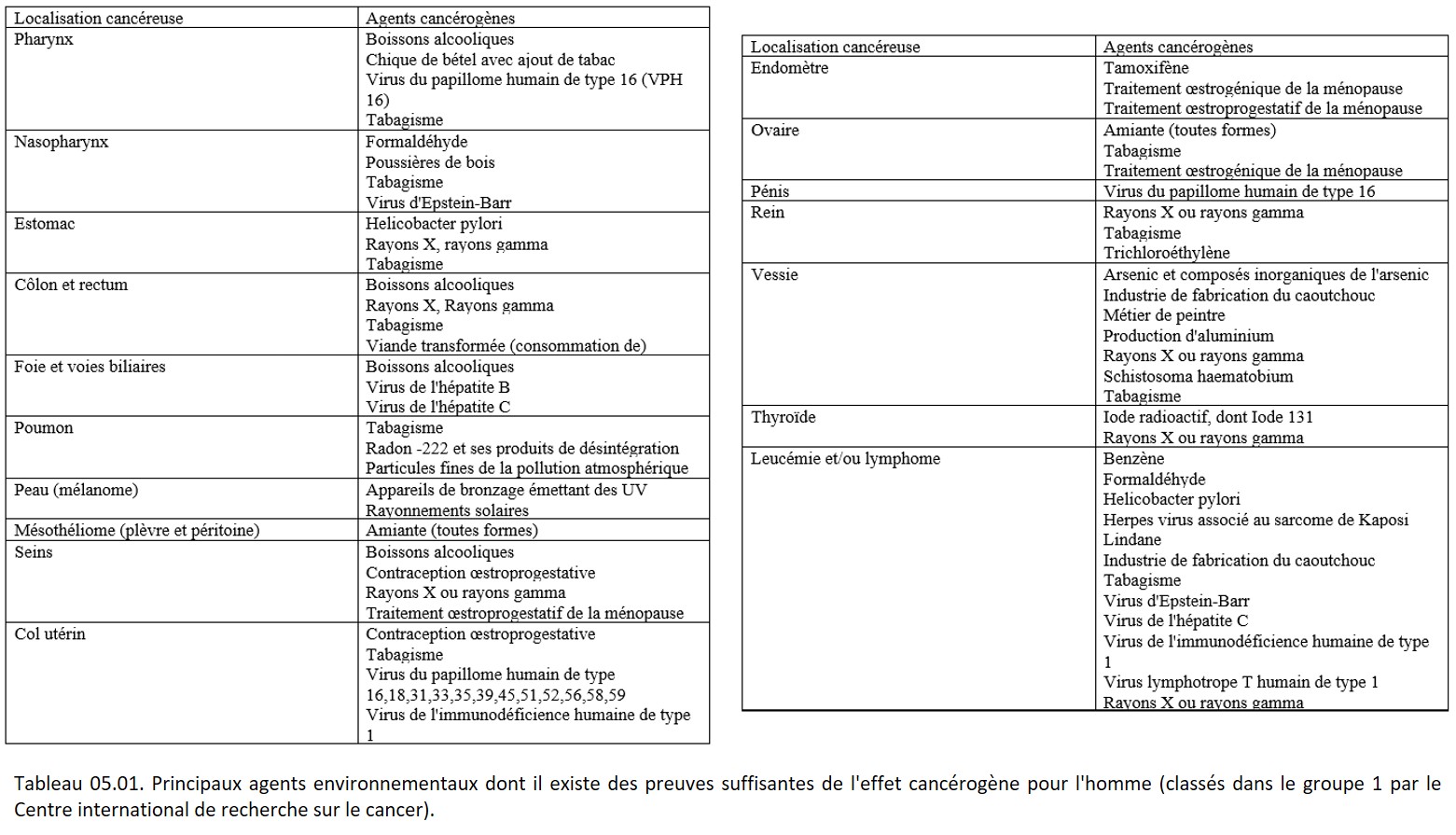{kind=link}
• carcinogènes chimiques : hydrocarbures polycycliques aromatiques (tabac), amines aromatiques (colorants, industrie du caoutchouc),
• virus (hépatite B, d'Epstein-Barr, etc.),
• radiations ;
• Agents promoteurs : ils favorisent l'expression d'une lésion génétique, préalablement induite par un agent initiateur. Ils n'induisent pas de lésions de l'ADN. Le temps écoulé entre l'initiation et l'apparition des tumeurs est réduit en présence d'agents promoteurs. Exemples (tableau 05.01) :
• hormones : œstrogènes (cancer du sein),
• nutrition : alcool (tumeurs ORL),
• schistosomiase et cancer de la vessie.
Trois familles de gènes impliquées dans la cancérogenèse
Oncogènes
Certains virus animaux sont capables d'induire des tumeurs (ex. : sarcome de Rous du poulet, découvert en 1911). Les propriétés transformantes de ces virus sont dues à la présence dans leur génome de séquences particulières, les oncogènes viraux (v-onc).
Ces gènes renferment à eux seuls toute l'information pour l'activité transformante. Ces gènes sont des formes altérées de gènes normaux d'origine cellulaire, les proto-oncogènes, capturés par les rétrovirus au cours de leur réplication.
Les proto-oncogènes sont présents dans toutes les espèces (de l'insecte à l'homme) et jouent un rôle essentiel dans des étapes clés de la régulation de l'embryogénèse ou de la croissance cellulaire ou tissulaire. Ces gènes normaux lorsqu'ils sont remaniés et/ou sur-exprimés deviennent des oncogènes (c-onc). Ils peuvent induire l'apparition et/ou le développement d'une tumeur.
Les oncogènes sont schématiquement classés en :
• gènes immortalisants (ex : c-myc) codant pour des protéines nucléaires se liant à l'ADN ;
• gènes transformants (ex. : KRAS, RET, KIT) (tableau 05.02).
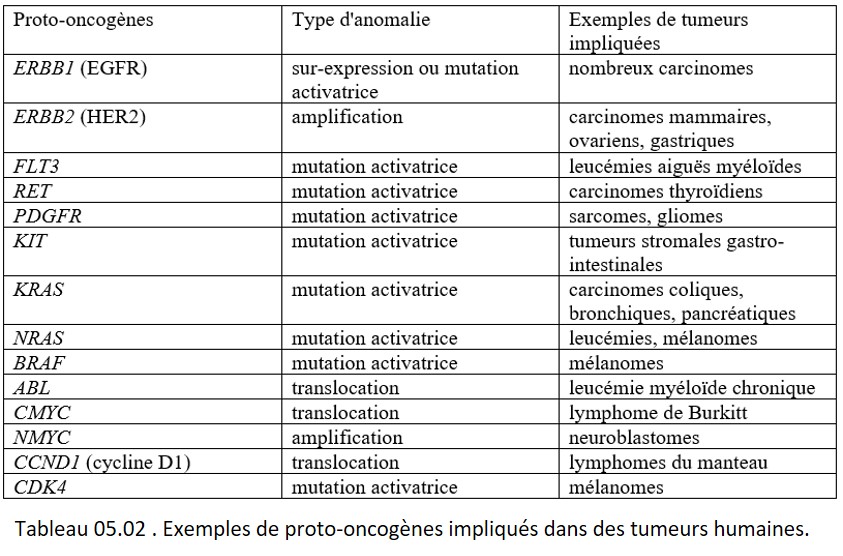{kind=link}
Gènes suppresseurs de tumeur
Les gènes suppresseurs de tumeur (ou anti-oncogènes) sont des inhibiteurs de la croissance cellulaire. L'inactivation biallélique de ces gènes se traduit par la perte de fonction de leur produit protéique.
Le premier gène suppresseur de tumeur décrit est le gène Rb du rétinoblastome. Le gène suppresseur de tumeur le plus souvent impliqué est TP53, avec des mutations somatiques dans de très nombreux cancers et des mutations germinales dans le syndrome de Li-Fraumeni.
Les oncogènes et gènes suppresseurs de tumeur codent pour des protéines qui interviennent dans les grandes fonctions cellulaires : signalisation, prolifération, différenciation, cycle, apoptose (tableau 05.03).
{kind=link}
Gènes de maintien de l'intégrité du génome (care takers)
Chaque réplication de l'ADN d'une cellule génère une multitude d'erreurs de copie de la séquence primaire et de cassure de l'ADN. De plus des agents pathogènes (rayons X, UV, hydrocarbures) peuvent entraîner des lésions ponctuelles de l'ADN (cassure d'un brin, délétion, mutation d'une base). Les gènes de maintien de l'intégrité codent pour des complexes multi-fonctionnels capables de surveiller l'intégrité du génome (exemple : MSH2, MSH6). En cas d'anomalies, différents systèmes de réparation sont mis en place (exemple : BRCA1, rad50, MLH1). S'ils échouent, la cellule lésée meurt par apoptose.
L'altération des deux allèles de ces gènes conduit à une susceptibilité accrue aux cancers, par instabilité génétique (accumulation de mutations conduisant à l'activation d'oncogènes ou à l'inactivation d'anti-oncogènes) ou chromosomique.
Des mutations impliquant ces trois familles de gènes sont présentes dans la majorité des cancers. Ces lésions peuvent être constitutionnelles (héritées par voie génétique) ou somatiques (d'origine environnementale).
Contrôle de l'expression et/ou de l'activation des gènes impliqués dans la cancérogenèse
Plusieurs mécanismes peuvent être responsables de l'expression et/ou de l'activation des gènes impliqués dans la tumorigenèse. Ces mécanismes ne sont pas mutuellement exclusifs.
Mutations ponctuelles, délétions, insertions
Pour les proto-oncogènes, un seul événement génétique est généralement suffisant pour l'activation (dominant). Pour les gènes suppresseurs de tumeurs et les gènes de maintien de l'intégrité du génome, un double événement est nécessaire pour que le gène soit inactivé au niveau des deux allèles (récessif) (figure 05.03)
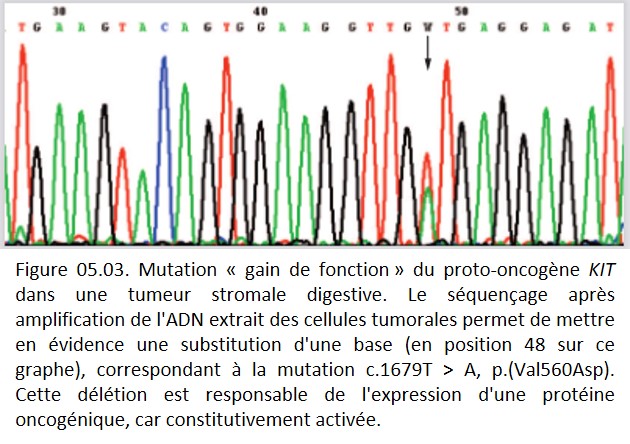{kind=link}
Amplification génique
Ce phénomène correspond à une multiplication du nombre de copies d'un gène. Il en résulte une augmentation de son expression. Il serait surtout tardif dans l'oncogenèse (figure 05.04).
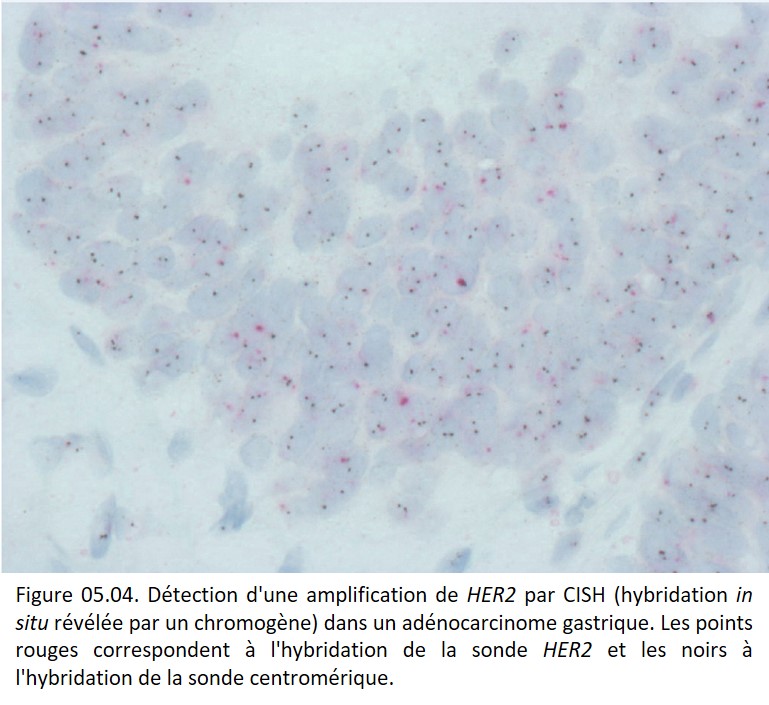{kind=link}
Réarrangements chromosomiques
Les translocations peuvent aboutir soit à l'expression d'une protéine chimérique résultant de la fusion entre deux gènes, soit à l'hyperexpression d'un oncogène en raison de la transposition de la région codante de celui-ci à proximité de séquences régulatrices d'autres gènes.
Exemple 1
Dans la leucémie myéloïde chronique (LMC) la translocation réciproque entre les chromosomes 9 et 22 produit un chromosome 22 raccourci : le chromosome de Philadelphie. Cette translocation aboutit à un gène de fusion BCR-ABL codant pour une tyrosine kinase activée. Il existe actuellement des molécules thérapeutiques capables de bloquer spécifiquement la tyrosine kinase activée par cette translocation. Grâce à cette thérapeutique (Imatinib, nilotinib), le pronostic de la LMC a été transformé.
Exemple 2
Dans le lymphome de Burkitt, la translocation (8 ;14) aboutit à la surexpression de l'oncogène CMYC (chromosome 8) qui se retrouve sous le contrôle du promoteur de la chaîne lourde des immunoglobulines (chromosome 14).
Délétions chromosomiques et remaniements chromosomiques complexes
Il peut en résulter une perte de fonction d'un gène suppresseur de tumeur. Cette perte de fonction dans la cellule peut être récessive (ex. : RB) ou dominante (ex. : P53).
Mécanismes épigénétiques
L'hypo- ou l'hyper-méthylation de gènes ou de leurs séquences régulatrices peut moduler leur transcription, alors que la séquence d'ADN est normale. On parle de mécanismes épigénétiques par opposition aux mécanismes génétiques c'est-à-dire avec altération de l'ADN.
Facteurs favorisant les altérations des gènes impliqués dans la cancérogenèse
Facteurs héréditaires
Ces facteurs génétiques sont responsables de prédispositions familiales aux cancers. La transmission peut être dominante ou récessive, et la pénétrance variable. Les formes héréditaires de cancer résultent essentiellement de l'inactivation constitutionnelle de gènes suppresseurs mais elles sont parfois secondaires à une activation constitutionnelle de proto-oncogènes ou à une inactivation constitutionnelle des gènes de réparation de l'ADN. Les prédispositions génétiques aux cancers sont nombreuses, et les prédispositions monogéniques sont les mieux connues (tableau 05.04). À ce jour, plus d'une centaine de gènes de prédisposition génétique aux cancers ont été identifiés.
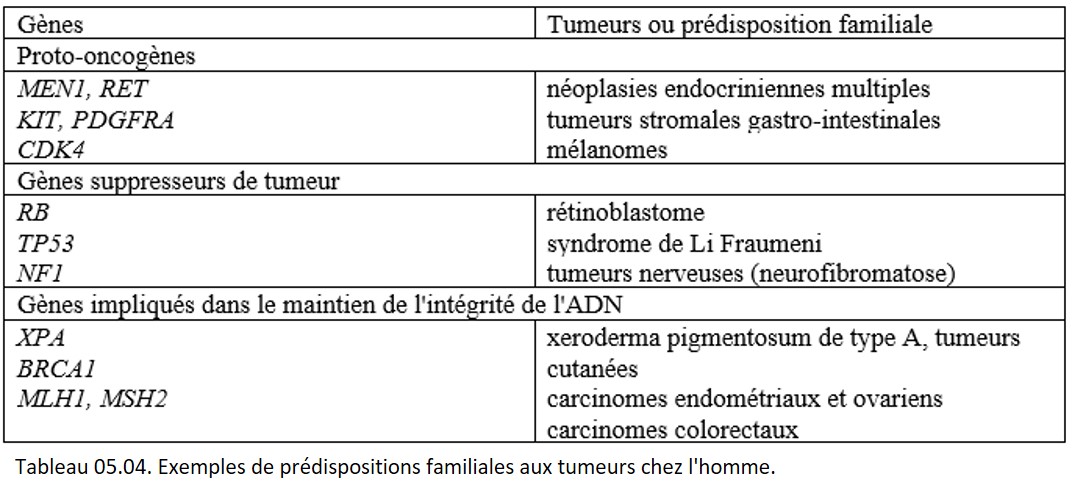{kind=link}
Facteurs viraux
• Rétrovirus à ARN. Certains rétrovirus sont directement oncogéniques, mais il n'en existe d'exemple connu que chez l'animal. Chez l'homme, le rétrovirus HTLV1 s'intègre au hasard dans le génome, il est dépourvu d'oncogène mais contient un gène transactivateur (tax) capable d'activer les gènes de l'interleukine 2 et de son récepteur dans les lymphocytes T.
• Virus oncogènes à ADN : ils ne renferment pas d'oncogène de type v-onc. Le plus souvent ils semblent agir par trans-activation de gènes cellulaires (mutagénèse insertionnelle).
• D'autres virus agissent de façon plus indirecte.
Exemple
Le virus d'Epstein-Barr induit chez les sujets immunodéprimés (VIH, endémie paludique, transplantés) une intense prolifération polyclonale des lymphocytes B infectés et augmente ainsi le risque de survenue de translocations chromosomiques. Au cours de ces translocations somatiques peuvent se produire des juxtapositions accidentelles de gènes, capables d'activer des proto-oncogènes : la translocation t(8 ;14) : juxtaposition de CMYC et du gène de la région constante des immuno-globulines.
Facteurs physiques
• Les radiations ionisantes favorisent les mutations et les cassures chromosomiques.
• Les ultraviolets entraînent des cassures de l'ADN, impossibles à réparer chez les malades atteints de xeroderma pigmentosum (anomalie génétique) ; d'où la survenue de multiples cancers cutanés.
Facteurs chimiques
Il en existe de nombreux exemples :
• le tabac ;
• l'aflatoxine induit des mutations très spécifiques du gène TP53 : carcinogène hépatique ;
• les cancers professionnels dus aux dérivés du benzopyrène.
Progression tumorale et cycle cellulaire
La progression du cycle cellulaire est finement régulée par des « points de contrôle », qui permettent notamment une régulation de la vitesse de prolifération et un maintien de l'intégrité du génome cellulaire. Dans beaucoup de tumeurs, ces points de contrôle sont altérés.
En cas de cancer, les signaux extra-cellulaires ou intracellulaires reçus par la cellule vont être capables d'activer les complexes cycline/CDK ou d'altérer l'activité des inhibiteurs (p21, p15, p16). Le résultat sera la levée du verrou Rb et l'entrée de la cellule en cycle.
Exemple
Le cancer du col de l'utérus : les papillomavirus humains (HPV) sont des petits virus à ADN double brin capables d'infecter les tissus épithéliaux, le plus souvent de façon asymptomatique. Certains types d'HPV dits à haut risque (HPV 16, 18) sont associés au cancer du col de l'utérus. On sait désormais que ce virus s'intègre dans le génome de la cellule hôte où il code pour des protéines virales (E6 et E7) capables de se lier et de dégrader respectivement p53 et Rb, ce qui entraîne une levée du verrou du cycle cellulaire.
Progression tumorale et apoptose
La cellule cancéreuse devient résistante à l'apoptose.
L'apoptose est impliquée dans le contrôle de l'homéostasie cellulaire, et est sous le contrôle de nombreux gènes :
• gènes pro-apoptotiques ; exemple : TP53, MYC (voir schéma du cycle cellulaire) ;
• gènes de survie ou anti-apoptotiques ; exemple : BCL2.
En cas de lésion de l'ADN, le gène TP53 est activé, permettant, par l'intermédiaire de p21, l'arrêt du cycle cellulaire et la réparation des lésions de l'ADN ou l'activation de l'apoptose. Il existe des anomalies du gène TP53 dans 2/3 des cancers (mutations, délétions) entraînant la suppression du point de vérification de G1 et donc de la voie apoptotique en cas d'instabilité génomique ou d'anomalies chromosomiques.
Dans le lymphome folliculaire, la translocation t(14 ;18) aboutit à la juxtaposition du gène BCL2 avec le locus de la chaîne lourde des immunoglobulines et entraîne la surexpression de la protéine BCL-2. L'accumulation de cette protéine anti-apoptotique augmente la survie des lymphocytes B, ce qui accroît le risque d'acquisition de nouvelles anomalies génétiques conduisant au développement du lymphome folliculaire.
Progression tumorale et immortalité : la cellule cancéreuse a une prolifération illimitée
Les cellules normales sont programmées pour un nombre limité de dédoublements (environ 60–70 in vitro). Aux extrémités des chromosomes se trouvent des séquences répétitives (télomères) qui sont érodées à chaque réplication de l'ADN. Leur disparition induit un arrêt de la prolifération (G0).
Dans la plupart des cellules tumorales, il existe un maintien des télomères au cours des réplications successives. Ceci est dû à la surexpression des télomérases, qui sont les enzymes capables d'ajouter des séquences répétées à l'extrémité des chromosomes.
Modifications fonctionnelles et morphologiques
Fiche signalétique de la cellule cancéreuse
D'un point de vue fonctionnel, on reconnaît aux cellules cancéreuses des propriétés communes qui les différencient des cellules normales :
1. indépendance vis-à-vis des signaux de prolifération (facteurs de croissance) provenant de l'environnement ;
2. insensibilité aux signaux anti-prolifératifs ;
3. résistance à l'apoptose ;
4. prolifération illimitée (perte de la sénescence) ;
5. capacité à induire l'angiogénèse ;
6. capacité d'invasion tissulaire et de diffusion métastatique.
Ces anomalies fonctionnelles sont l'aboutissement d'un processus multi-étapes dans lequel l'environnement n'est pas neutre. Elles s'accompagnent de modifications morphologiques de la cellule qui permettent le plus souvent de reconnaître son caractère cancéreux en l'observant au microscope optique.
Il faut cependant faire deux remarques :
• aucune de ces anomalies morphologiques prises séparément n'est spécifique de la cellule cancéreuse (en dehors pour certains auteurs des figures de mitoses anormales) ;
• certaines tumeurs au comportement authentiquement malin sont constituées de cellules morphologiquement très proches de leur contrepartie normale ; d'autres critères morphologiques (mauvaise limitation, invasion vasculaire) ou évolutifs (métastases) sont alors nécessaires pour affirmer la malignité.
Modifications du noyau
Noyau en mitose
• Augmentation du nombre de cellules en mitoses.
• Mitoses anormales (figure 05.05).
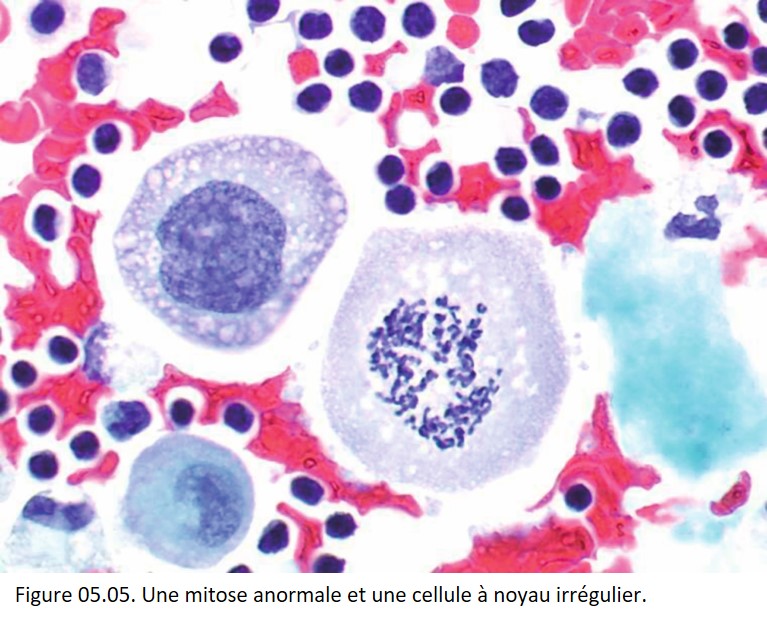{kind=link}
Noyau interphasique
• Anisocaryose (du grec aniso = différent et caryo = noyau) : inégalité de taille d'un noyau à l'autre.
• Augmentation du rapport nucléo-cytoplasmique : le plus souvent due à une augmentation de la taille du noyau.
• Hyperchromatisme : aspect dense et sombre du noyau lié à une condensation ou à une augmentation du nombre des chromosomes (aneuploïdie).
• Irrégularités de forme et de contours (figures 05.06, 05.07, 05.08, 05.09 et 05.10).
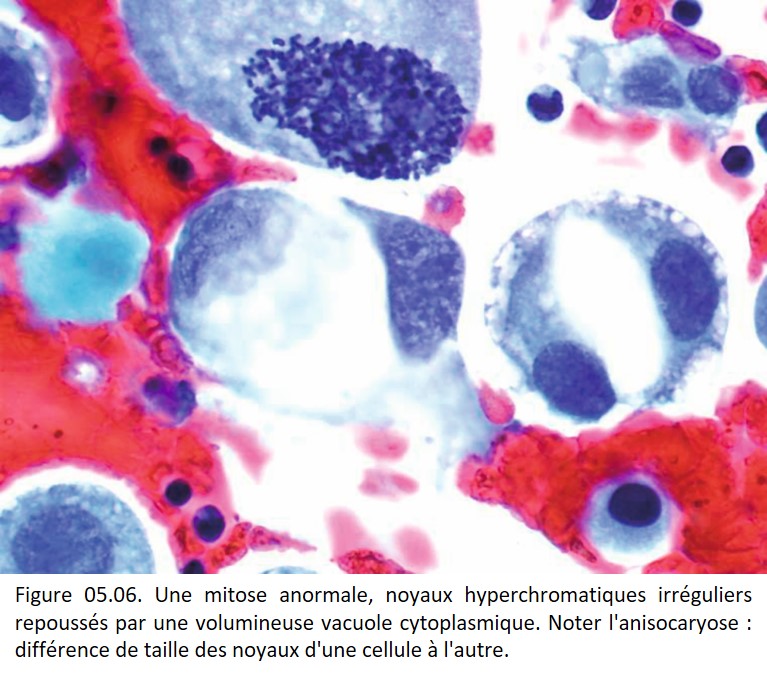{kind=link}
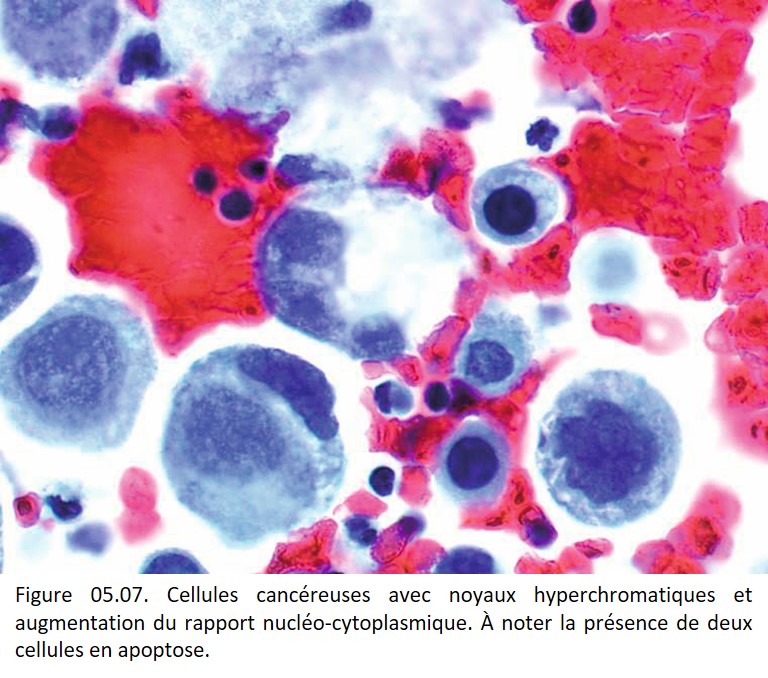{kind=link}
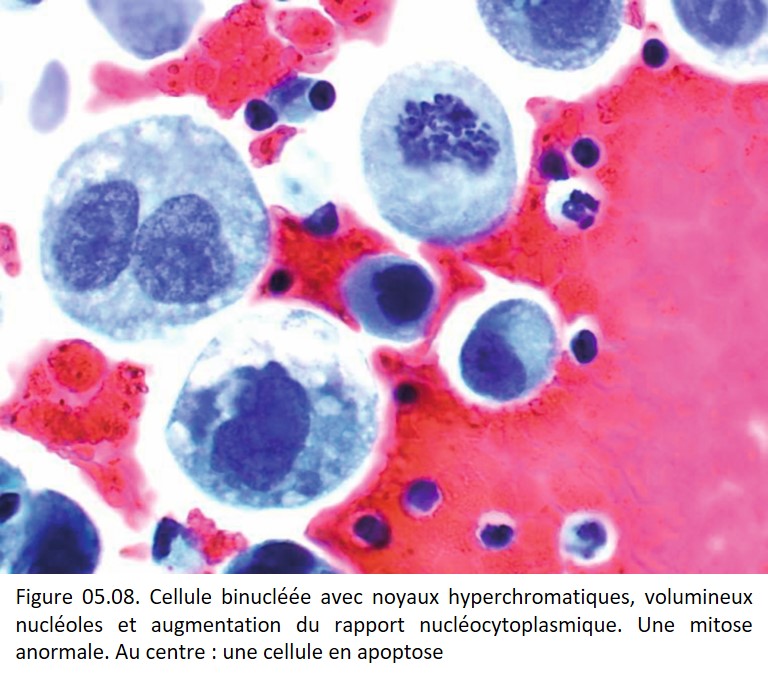{kind=link}
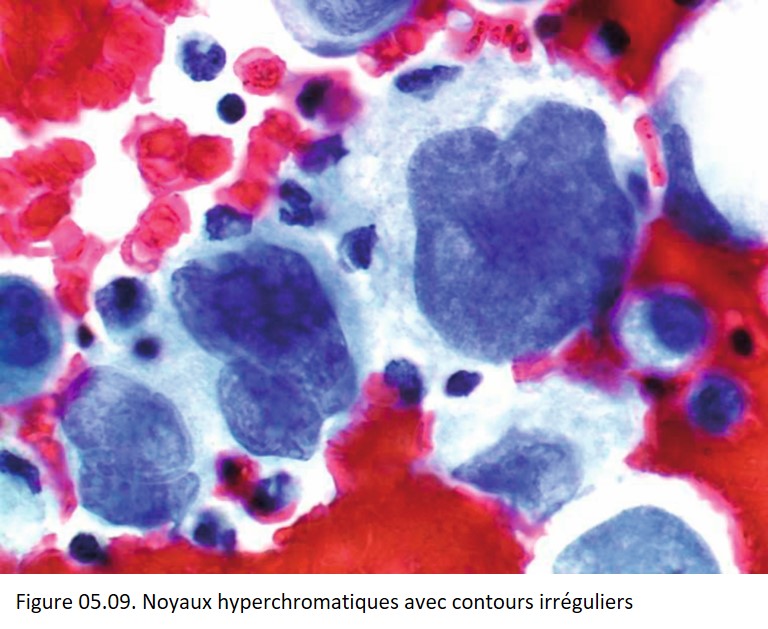{kind=link}
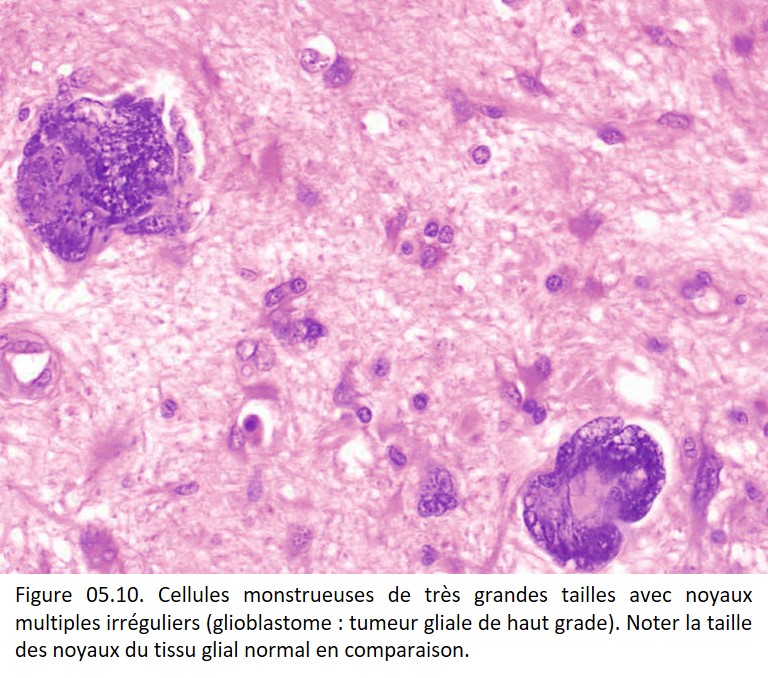{kind=link}
• Multinucléation (figure 05.09).
Modifications du cytoplasme
Cytosquelette
Dans la cellule normale, le cytosquelette est constitué de trois types de filaments :
1. microtubules : structures de 20–25 nm d'épaisseur constituées principalement de polymères de tubulines ;
2. microfilaments : structures contractiles de 6–8 nm d'épaisseur contenant notamment des filaments d'actine ;
3. filaments intermédiaires : les plus importants sont les filaments de cytokératine (présents dans les cellules épithéliales et mésothéliales) et de vimentine (surtout dans les cellules conjonctives = mésenchymateuses).
Dans la cellule cancéreuse, le cytosquelette est le plus souvent conservé, avec des anomalies de répartition. Il n'est pas visible en microscopie optique mais ses constituants peuvent être mis en évidence par immunohistochimie. Cette conservation est intéressante pour le pathologiste car la mise en évidence de tels ou tels types de filaments intermédiaires par exemple, aide à préciser le tissu d'origine d'une cellule cancéreuse.
Système sécrétoire
• Variations visibles sur les colorations standards, telles des vacuoles cytoplasmiques (excès de mucus) refoulant le noyau dans les adénocarcinomes mucosécrétants, ou un cytoplasme clair, optiquement vide (accumulation anormale de glycogène) dans les cancers du rein à cellules claires par exemple (figure 05.11).
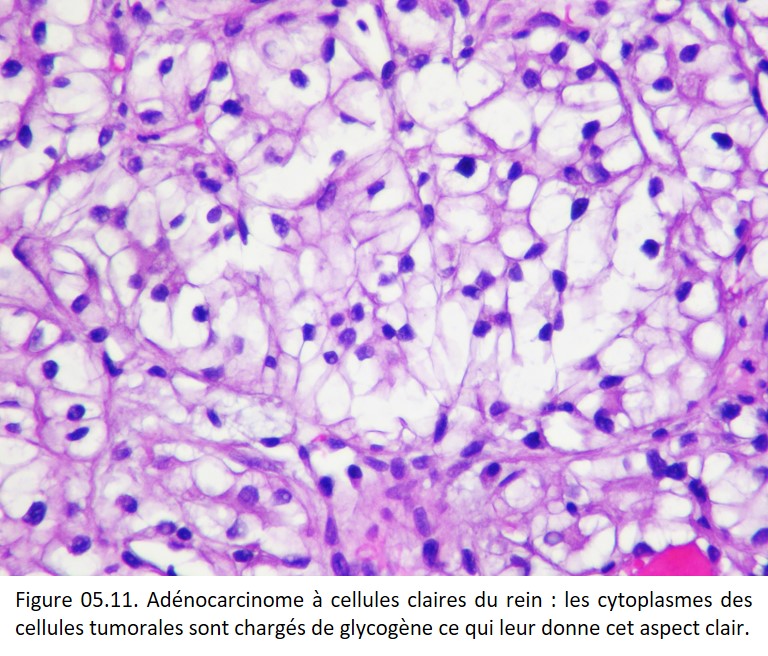{kind=link}
• Variations quantitatives des sécrétions normales (ex. : pic d'immunoglobulines monoclonales dans le myélome).
• Apparition de substances nouvelles, soit par dérépression d'une synthèse de protéines de type fœtal (ex : alpha fœtoprotéine, antigène carcino-embryonnaire = ACE), soit par sécrétion inappropriée d'une hormone (ex. : sécrétion d'ACTH par certains carcinomes à petites cellules du poumon). Ces substances, considérées comme des marqueurs tumoraux, peuvent être dosées dans le sérum lorsqu'elles sont sécrétées ou identifiées in situ par immuno-histochimie.
Membrane
La membrane joue un rôle crucial dans les échanges entre les cellules et les interactions avec le milieu extra-cellulaire.
Aspects morphologiques
Les modifications morphologiques ne sont visibles qu'en microscopie électronique : irrégularités, microvillosités, bulles, projections, modifications des systèmes de jonction. Elles ne sont pas prises en compte pour le diagnostic de cancer en routine.
Il existe des modifications des protéines de surface, et notamment des molécules d'adhérence, qui sont impliquées dans les interactions intercellulaires et cellule-matrice extra-cellulaire.
Aspects fonctionnels
• Anomalies des récepteurs membranaires : augmentation de nombre et perte de régulation.
• Modifications des enzymes membranaires : augmentation des enzymes protéolytiques (protéases, glycosidases) favorisant la dégradation de la substance intercellulaire.
• Modifications des antigènes de membrane :
altération ou perte des antigènes normaux (Ag d'espèces, d'organes ou de tissus),
apparition de néoantigènes : ré-expression d'antigènes embryonnaires : alpha fœtoprotéine, antigène carcinoembryonnaire,
expression anormale d'antigène de différenciation, d'Ag associés aux virus (ex : protéine latente de membrane du virus d'Epstein-Barr virus ; antigènes viraux EBNA1, LMP1 et LMP2 dans le lymphome de Hodgkin).
• Modifications de la perméabilité membranaire : l'augmentation de perméabilité pour différents cations (Ca++ et Mg++) joue un rôle dans plusieurs fonctions cellulaires, en particulier la prolifération.
Stroma tumoral
Le stroma tumoral est caractérisé par tout ce qui est présent au sein d'une tumeur et n'est pas une cellule tumorale. Le stroma comprend donc notamment le tissu conjonctif, les vaisseaux et les leucocytes. Le stroma sert de charpente à la tumeur et assure ses apports nutritifs. Il est sous la dépendance du tissu tumoral dont les cellules peuvent, par exemple, élaborer des substances qui vont favoriser la pousse des vaisseaux. Il est d'usage de réserver le terme de stroma au support conjonctif des tumeurs malignes et de ne pratiquement pas l'utiliser dans le cas des tumeurs bénignes, mais rien ne s'y opposerait conceptuellement.
C'est dans les carcinomes invasifs que le stroma est le plus nettement individualisé. Il y a cependant un stroma dans toutes les autres tumeurs solides, constitué au minimum des vaisseaux et d'une matrice extra-cellulaire d'abondance variable.
Les variations morphologiques du stroma sont multiples, certaines d'entre elles sont caractéristiques d'un type tumoral donné et auront donc une valeur séméiologique pour le diagnostic du type tumoral (figure 05.12).
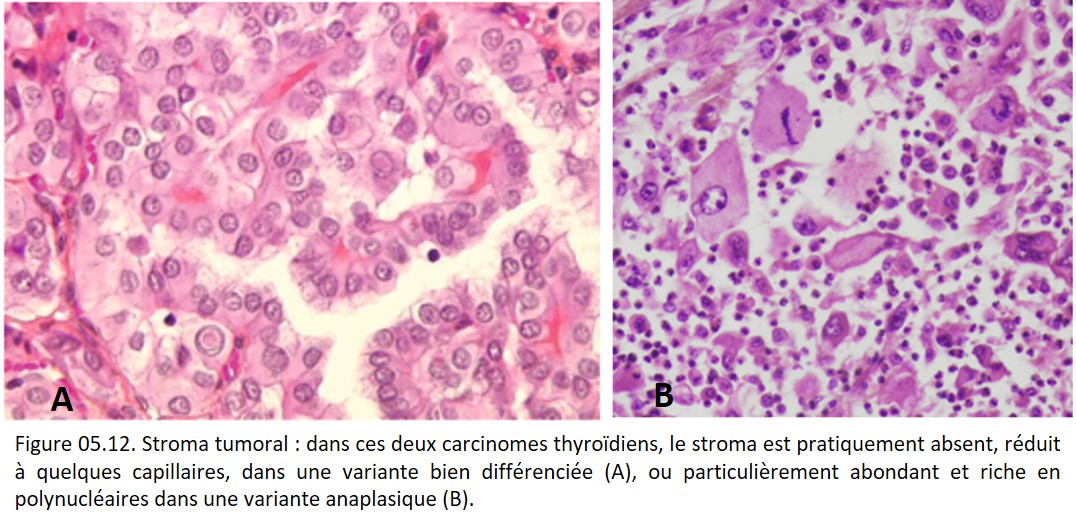{kind=link}
Variations quantitatives
Certains carcinomes très différenciés ont un stroma qui peut être exactement proportionné à la prolifération épithéliale. Dans les tumeurs endocriniennes, le stroma comporte souvent des capillaires sinusoïdes semblables à ceux d'une glande endocrine normale (stroma adaptatif).
Plus souvent, le stroma est disproportionné par rapport à la prolifération épithéliale :
• lorsqu'il est relativement peu abondant, la tumeur sera molle, souvent nécrosée, semblable macroscopiquement à du tissu cérébral. C'est un cancer que l'on caractérisera macroscopiquement d'« encéphaloïde » ;
• à l'inverse, lorsqu'il est très abondant, riche en fibres collagènes, la tumeur sera dure et rétractée, c'est le squirrhe. Cette rétraction, comparable à celle de certaines cicatrices pathologiques, est liée à la présence de nombreux myofibroblastes.
Variations qualitatives
Le tissu conjonctif du stroma possède certaines propriétés réactionnelles du tissu conjonctif normal. Il peut s'y produire une réaction inflammatoire. Celle-ci surviendra, par exemple, lors de la destruction du tissu tumoral par une irradiation. La nécrose des cellules tumorales déclenche une réaction exsudative. Il peut même se produire une réaction à corps étrangers autour de squames de kératine élaborées par la tumeur. Dans certaines tumeurs, la réaction inflammatoire du stroma est une réaction tuberculoïde.
Quelques tumeurs ont un stroma riche en cellules lymphocytaires ou plasmocytaires, ce qui peut être la manifestation d'une réaction immunitaire (ex. séminome testiculaire, thymome type B1, carcinome lympho-épithélial). Cet aspect va parfois de pair avec un pronostic meilleur.
Cancer et angiogenèse
La néovascularisation issue de l'angiogenèse tumorale présente un état d'activation cellulaire maximum pour une efficacité de perfusion médiocre. Elle est très hétérogène en densité, par sa maturation phénotypique d'une zone tumorale à l'autre et d'une tumeur à l'autre.
Une tumeur ne peut pas croître au-delà de 1 à 2 mm sans l'aide d'une riche vascularisation sanguine. Les rapports entre le tissu tumoral proprement dit et sa vascularisation sont donc critiques dans l'histoire naturelle de chaque cancer.
La vasculogenèse est une prolifération vasculaire due à la différenciation de cellules précurseurs, communes aux lignées sanguines, en cellules endothéliales qui se répandent, s'associent et établissent un réseau vasculaire. Ce terme est très majoritairement réservé aux étapes correspondantes de l'embryogenèse.
L'angiogenèse est une prolifération vasculaire due au bourgeonnement vasculaire à partir de vaisseaux préexistants, puis à l'installation d'un réseau et à sa différenciation en différents secteurs fonctionnels. Ce processus implique le recrutement et la différenciation de cellules péricytaires et de cellules musculaires lisses, qui concourent à stabiliser le nouveau réseau et à lui donner une efficacité fonctionnelle. L'angiogenèse est souvent liée aux processus inflammatoires ou tumoraux.
Vascularisation en périphérie des tumeurs
Dans la zone périphérique d'invasion tumorale, la prolifération des cellules endothéliales est active et elle produit de nouveaux vaisseaux souvent anormaux. La prolifération vasculaire est particulièrement vigoureuse et l'index de prolifération des cellules endothéliales est 50 à 200 fois plus élevé que pour les mêmes cellules des tissus normaux.
Les vaisseaux créés au sein de la tumeur sont anormaux. Ce sont des canaux à paroi mince plutôt de type veinulaire, irrégulièrement anastomosés avec de nombreux culs-de-sac. Ils ont tendance à former des shunts artério-veineux. La bordure endothéliale est incomplète (sauf dans les tumeurs cérébrales primitives), la membrane basale est souvent absente, les cellules satellites (péricytes et cellules musculaires lisses) raréfiées. Il n'y a pas d'innervation et de nombreux espaces vasculaires sont bordés directement par les cellules tumorales.
Ces néo-vaisseaux ne sont généralement que peu contrôlables par les mécanismes locaux habituels (mécanisme nerveux et système des cytokines) que l'on observe par exemple au cours de l'inflammation. L'efficacité de perfusion est médiocre. Les courts-circuits artério-veineux s'opposent à une perfusion capillaire efficace. Le régime liquidien est chaotique avec des inversions de flux et une stase selon une période de 2–3 min. Le drainage des fluides interstitiels est souvent déficient en liaison avec l'excès de perméabilité et l'absence de drainage lymphatique fonctionnel. Enfin, cette vascularisation est très inégalement répartie en densité d'un secteur à un autre de la tumeur.
Dans la zone périphérique d'invasion tumorale, on retrouve des taux élevés de facteur de croissance endothélial vasculaire (VEGF), du facteur de croissance fibroblastique basique (FGFb), de la phosphorylase de la thymidine (TP). Tous ces facteurs sont induits par l'hypoxie.
Vascularisation au centre des tumeurs
Au fil de la croissance tumorale, les marges s'incorporent dans le centre de la tumeur, mêlant néovascularisation et vascularisation d'origine de l'hôte. La densité de microcirculation devient 4 à 10 fois plus faible qu'au niveau des berges. Les cellules tumorales s'adaptent à l'hypoxie en activant la glycolyse anaérobie. Les cellules endothéliales activent la fabrication des molécules du stress hypoxique (VEGF, TP, complexe VEGF/récepteur du VEGF) et les inhibiteurs de l'apoptose (BCL-2). Quand le mécanisme anti-apoptotique endothélial défaille, les cellules tumorales sont en situation d'accès facile au compartiment intravasculaire.
Immunité anti-tumorale
Rôle du système immunitaire dans le contrôle tumoral
La réponse immune anti-tumorale joue un rôle majeur dans la défense de l'organisme contre les tumeurs. Elle fait intervenir :
• l'immunité innée, avec notamment des cellules cytotoxiques (ex : lymphocytes natural killer (NK), macrophages, neutrophiles), et des facteurs solubles (ex. : interféron gamma), qui peuvent avoir des effets directs ou indirects (pro-inflammatoire ou anti-angiogénique). Ces cellules sont capables de détruire les cellules tumorales de façon non spécifique. Elles sont la première réponse immunitaire, rapide et immédiate, permettant d'orienter la réponse adaptative ;
• l'immunité adaptative, c'est-à-dire dépendante de la reconnaissance des antigènes produits par la tumeur. La plupart des cellules tumorales expriment des antigènes de spécificité et d'immunogénicité variable. La présentation de ces antigènes par les cellules de l'immunité innée permet l'activation d'une réponse spécifique et la mise en place de mécanismes effecteurs.
Les mécanismes effecteurs de la réponse immune anti-tumorale sont :
• la cytotoxicité directe par les lymphocytes NK, les lymphocytes T cytotoxiques (CD8), ou les cellules dendritiques IKDC (Interferon gamma producing killer dendritic cells) ;
• la cytotoxicité médiée par les anticorps, qui paraît notamment très utile en thérapeutique, avec l'utilisation d'ac monoclonaux spécifiques de certains antigènes exprimés par les tumeurs (CD20, EGFRs) ;
• la production de facteurs solubles capables de moduler la réponse inflammatoire locale et/ou l'angiogénèse, tel l'interféron gamma.
Les étapes clefs impliquées dans une réponse immune anti-tumorale efficace ont été décrites et illustrées par Chen et Mellman, constituant « le cycle de l'immunité contre le cancer » (figure 05.13).
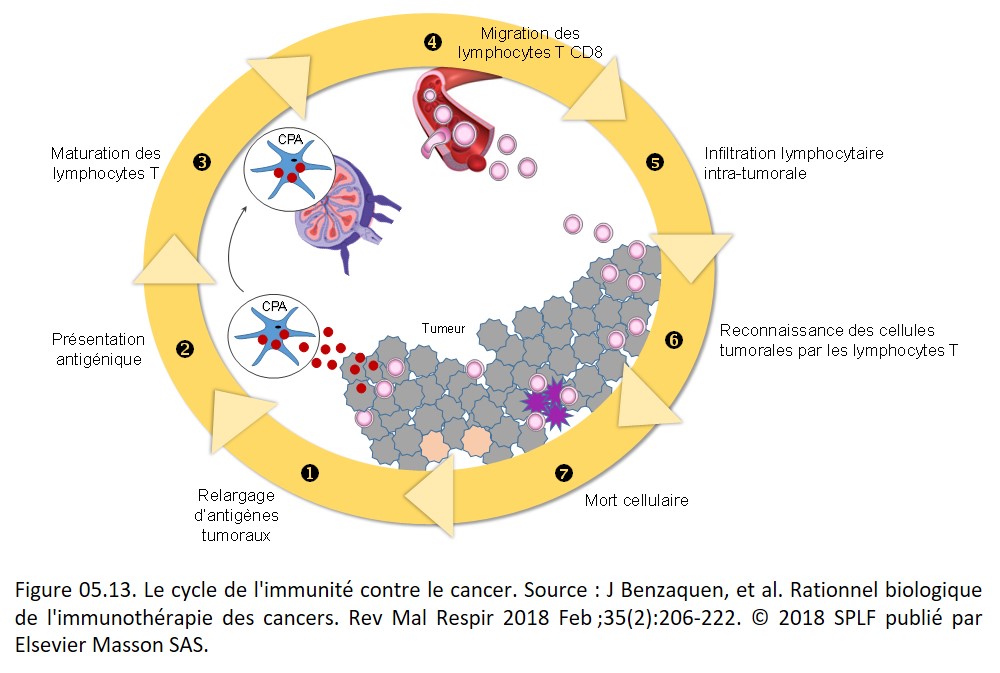{kind=link}
Les antigènes tumoraux qui apparaissent pendant le processus d'oncogenèse sont capturés par les cellules dendritiques du microenvironnement tumoral. Suite à leur activation, ces cellules dendritiques migrent via les vaisseaux lymphatiques vers les ganglions lymphatiques qui drainent la tumeur. C'est dans ces ganglions qu'elles présentent les antigènes tumoraux aux lymphocytes T ; ce qui conduit à leur activation et à leur différenciation en lymphocytes T effecteurs. Ces lymphocytes T effecteurs quittent les ganglions lymphatiques via les vaisseaux lymphatiques efférents pour se retrouver dans la circulation sanguine, puis dans les vaisseaux sanguins irrigants les tumeurs. Ils peuvent ainsi infiltrer le lit tumoral au travers de multiples processus impliquant des interactions intriquées entre les lymphocytes T et les cellules endothéliales, puis migrent dans la tumeur par voie trans-endothéliale. Une fois que les lymphocytes T anti-tumoraux ont infiltré le lit tumoral, ils peuvent reconnaître et détruire les cellules tumorales, ce qui a pour conséquence le relargage d'antigènes tumoraux supplémentaires et l'amplification de la réponse immunitaire. La lyse des cellules tumorales par les lymphocytes T anti-tumoraux met en jeu plusieurs mécanismes moléculaires, parmi lesquels :
• la libération de molécules cytotoxiques (perforine, granzyme, etc.) à partir de granules préformés ;
• l'activation des voies de signalisation en aval des récepteurs des molécules de la famille du TNF et notamment Fas et Trail.
La réponse immune anti-tumorale est probablement responsable du contrôle de la croissance tumorale dans une grande majorité de tumeurs, en particulier lors de la phase initiale de tumorigenèse.
Cependant, sous l'effet des mutations génétiques liées à l'instabilité génomique, et des modifications épigénétiques de l'ADN et des histones, les cellules tumorales, dont l'origine est monoclonale, se diversifient en sous-populations différentes selon leur capacité invasive, leur tendance à générer des métastases, leur sensibilité ou leur résistance au traitement, mais aussi dans leur capacité à induire une réponse immunitaire et leur sensibilité à cette réponse.
L'immunosurveillance peut moduler (« éditer ») l'immunogénicité des tumeurs en supprimant sélectivement les cellules exprimant fortement les antigènes reconnus par les lymphocytes T et de ce fait les plus sensibles à la réponse immunitaire, et en sélectionnant les clones cellulaires les plus résistants à l'immunosurveillance.
Les relations complexes entre l'immunosurveillance et l'échappement des cellules tumorales à la destruction médiée par le système immunitaire peuvent aboutir à trois scénarios constituant les 3 « E » de l'immunosurveillance : Élimination – Équilibre – Échappement.
La première phase est celle de l'élimination des cellules tumorales. Cette destruction survient généralement à un stade précoce de l'oncogenèse, avant que la tumeur ne soit détectable cliniquement. Parmi les exemples classiques de régression complète spontanée de tumeur primitive, on peut citer le « halo nevus » et le « burn out seminoma ». La seconde phase est l'instauration d'un équilibre entre le renouvellement des cellules tumorales et leur élimination par le système immunitaire. Cette étape peut se prolonger pendant plusieurs années de latence, durant lesquelles la tumeur peut être contrôlée et maintenue à son site d'origine. Mais cet équilibre peut être aboli par une évolution des clones cellulaires du fait de la plasticité de leur génome, conférant une résistance progressive envers l'action suppressive du système immunitaire ou par une défaillance des mécanismes d'immunosurveillance.
La troisième phase est représentée par l'échappement des cellules tumorales qui s'évadent du contrôle de l'immunosurveillance, se multiplient et envahissent les tissus environnants ou disséminent à distance. Cette étape induit un climat d'immunotolérance favorable à la croissance tumorale. La tumorigenèse est alors enclenchée, et la tumeur devient détectable et symptomatique cliniquement.
Échappement des tumeurs à la réponse immune
Les mécanismes d'échappement des tumeurs concernent à la fois la réponse immune innée et adaptative. Il peut s'agir :
• d'une immuno-sélection : sélection au cours du temps des sous-clones tumoraux ayant acquis des mécanismes d'échappement à la réponse immune. Ces sous-clones sont généralement sélectionnés en raison de la diminution de l'expression de cibles ou l'augmentation de l'expression d'inhibiteurs ;
• d'une immuno-subversion (induction d'une tolérance spécifique) mettant en jeu des phénomènes plus complexes de coopération intercellulaire.
Composition cellulaire d'une tumeur
Le tissu tumoral est constitué :
• de cellules tumorales : cellules prolifératives anormales ;
• d'un tissu de soutien (= stroma tumoral) fait de cellules et de substance extra-cellulaire dans laquelle est située la vascularisation tumorale. Les cellules du stroma ne contiennent pas les anomalies génétiques des cellules tumorales.
Types histologiques des tumeurs
Les différentes composantes de chaque tumeur (cellules tumorales et stroma) peuvent présenter des aspects morphologiques particuliers qui peuvent être regroupés par types histologiques. Les tumeurs sont ainsi classées en fonction de critères histologiques communs, définis par les classifications internationales, éditées par l'Organisation mondiale de la Santé (OMS), et remises à jour régulièrement. La reconnaissance de ces critères et le classement de la tumeur dans le type adéquat sont la base du diagnostic anatomopathologique de toute tumeur.
En pratique, les tumeurs sont classées en fonction de l'organe dont elles dérivent (ex. : sein, foie, rein, os) (figure 05.14), puis en fonction de leur type histologique. Le type histologique correspond à la cellule normale dont la tumeur semble dériver.
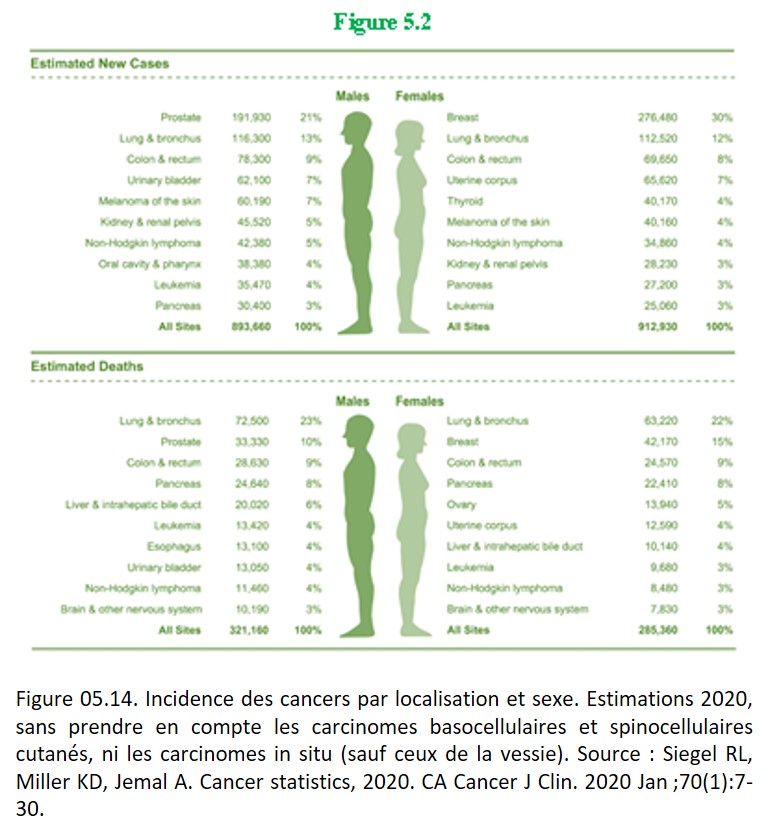{kind=link}
Au sein d'un même organe, les différents types histologiques de tumeur peuvent avoir une évolutivité particulière, un mode d'extension préférentiel et une réponse variable suivant les traitements. Les traitements sont donc adaptés en fonction du type histologique de chaque tumeur. Ainsi, bien que dans certains organes un type de tumeur soit nettement prépondérant (ex : les adénocarcinomes représentent 95 % des tumeurs malignes du sein), un autre type histologique de tumeur (ex. : un lymphome ou un angiosarcome) peut s'y développer et nécessitera un traitement radicalement différent. De plus, certains types de tumeurs peuvent être héréditairement transmissibles et justifier une enquête génétique, d'autres sont des maladies professionnelles (ex. : exposition à l'amiante et mésothéliome).
Différenciation tumorale
Le tissu tumoral tend à reproduire la structure et la fonction d'un tissu normal :
• soit le plus souvent, l'aspect du tissu dont les cellules tumorales sont originaires ;
• soit plus rarement un tissu différent : la tumeur est dite métaplasique.
Exemple : l'épithélium bronchique est bordé d'un épithélium cylindrique cilié. Les tumeurs bronchiques peuvent être des tumeurs à différenciation glandulaire ou des tumeurs à différenciation malpighienne lorsqu'elles surviennent sur une métaplasie malpighienne de l'épithélium bronchique.
La différenciation d'une tumeur est sa tendance à ressembler à un tissu normal ou embryonnaire. La tumeur est dite :
• bien différenciée, lorsqu'elle ressemble nettement et de façon homogène au tissu normal ;
• peu ou modérément différenciée lorsque la ressemblance est lointaine ou focale ;
• indifférenciée, ou anaplasique (ex. : carcinome indifférencié défini comme une tumeur à différenciation épithéliale dont il est impossible de préciser la différenciation glandulaire ou malpighienne) (figure 05.15).
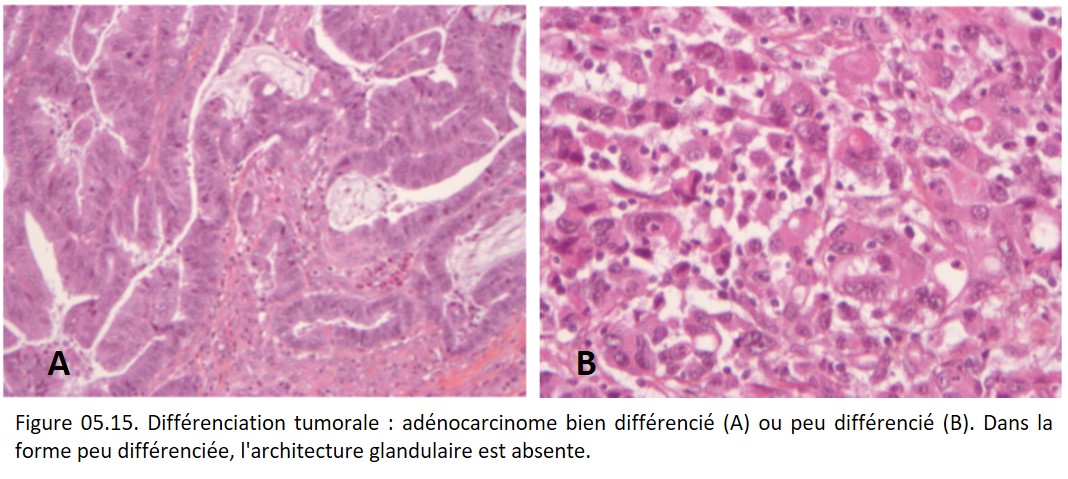{kind=link}
Tumeurs bénignes et malignes
Contrairement aux tumeurs bénignes, les tumeurs malignes aboutissent spontanément à la mort du patient. Cette distinction importante sur le plan évolutif est fortement corrélée à des critères macroscopiques et histologiques.
Tumeurs bénignes
Caractères évolutifs
Les tumeurs bénignes se développent localement et restent cantonnées au tissu dans lequel elles ont pris naissance. Leur croissance est lente. Toutefois, elles peuvent atteindre un volume et un poids importants. Elles ne récidivent pas après ablation chirurgicale, à condition que l'exérèse soit complète. Ces tumeurs ne métastasent jamais. Leur évolution est généralement favorable. Toutefois, dans certains cas, elles peuvent être la cause de complications graves voire mortelles, en raison de leur siège ou de désordres métaboliques.
Exemples :
• un méningiome du trou occipital, situé dans un orifice non expansible, peut avoir une évolution mortelle en provoquant un engagement du tronc cérébral à travers l'orifice occipital ;
• un adénome parathyroïdien est responsable d'une hyperparathyroïdie et par conséquent d'une hypercalcémie parfois dangereuse.
Caractères macroscopiques
Il s'agit de tumeurs circonscrites, bien limitées, nettement séparées des tissus avoisinants, parfois même entourées par une capsule (coque faite de tissu conjonctif). Cette limitation explique la facilité de l'exérèse chirurgicale et la possibilité d'une exérèse limitée à la seule tumeur (ex. : adénofibrome du sein, léiomyome de l'utérus).
Caractères histologiques
Le tissu tumoral reproduit de très près la structure du tissu initial (tumeur bien différenciée). Les cellules ont une morphologie normale et ne présentent aucun caractère de malignité (figure 05.16).
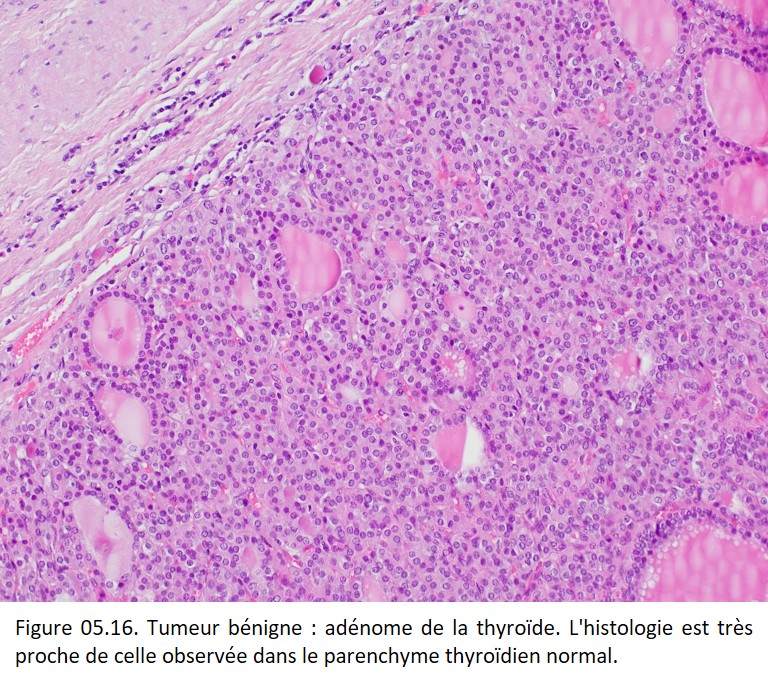{kind=link}
Il n'y a pas d'envahissement des tissus voisins. Les tumeurs bénignes refoulent sans les détruire les tissus sains de voisinage : elles sont expansives (ex. : adénome du foie).
Tumeurs malignes
Les caractères des tumeurs malignes ou cancers s'opposent point par point à ceux des tumeurs bénignes (tableau 05.05).
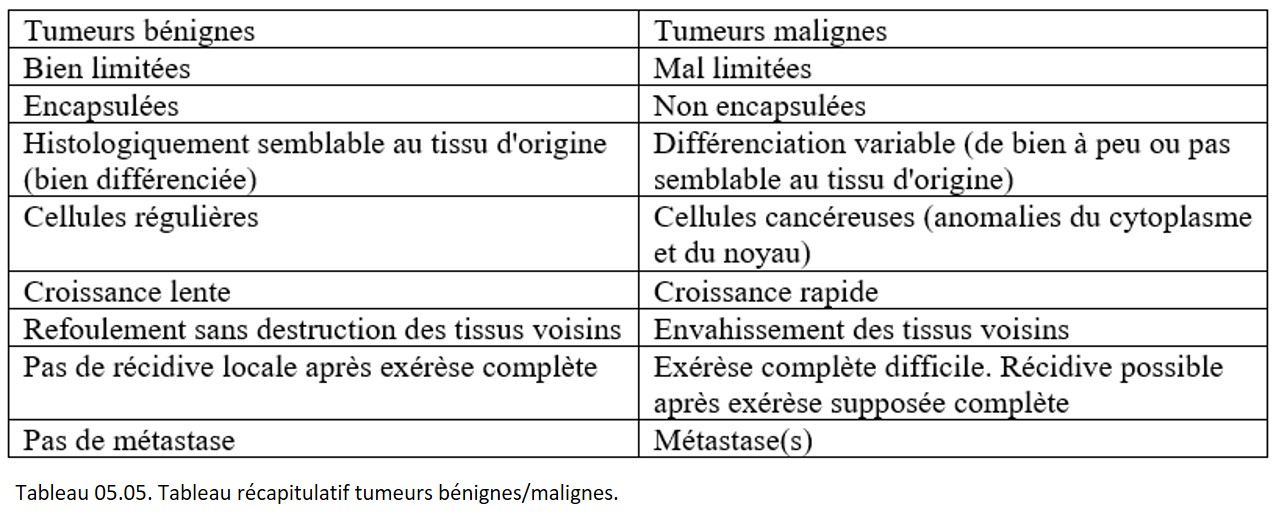{kind=link}
Caractères évolutifs
Les tumeurs malignes ont habituellement une croissance rapide. Elles donnent naissance à une dissémination tumorale à distance (surtout par voie lymphatique et sanguine) avec éclosion et développement de tumeurs secondaires dans d'autres viscères : les métastases. Les tumeurs malignes ont tendance à récidiver après éradication locale. L'évolution, en l'absence de traitement, se fait spontanément vers la mort.
Caractères macroscopiques
Les tumeurs malignes sont mal limitées, non encapsulées ; elles détruisent et envahissent l'organe dans lequel elles ont pris naissance, ainsi que les organes de voisinage. Leurs contours sont irréguliers. Les foyers de nécrose et d'hémorragie sont habituels.
Caractères histologiques
Les cellules tumorales malignes présentent habituellement des caractères anormaux (caractères cytologiques de malignité). Le tissu tumoral est plus ou moins différencié. Il « caricature » le tissu normal orthologue.
Limites de la distinction bénin/malin
Les caractères opposant les tumeurs bénignes et les tumeurs malignes constituent un schéma valable dans la plupart des cas. Toutefois, il est des cas où les critères morphologiques ne correspondent pas à l'évolution.
Les critères macroscopiques et microscopiques d'une tumeur ne permettent parfois pas d'en affirmer la nature bénigne ou maligne (ex. : tumeurs endocrines bien différenciées). Dans certains cas, cette nature maligne ne peut être affirmée que par la survenue de métastases (ex. : phéochromocytome).
Continuum entre certaines tumeurs bénignes et tumeurs malignes
Ce continuum appelé « progression tumorale » correspond à l'acquisition progressive par la tumeur bénigne d'un phénotype de malignité, d'anomalies chromosomiques et géniques en nombre croissant. L'inverse est impossible. Un exemple très fréquent est celui des adénomes coliques (bénins) qui peuvent se transformer en adénocarcinomes (malins). D'autres exemples incluent les tumeurs urothéliales papillaires, les astrocytomes, les tumeurs papillaires du rein.
Tumeurs d'agressivité locale
De rares tumeurs bénignes ont une forte tendance à l'envahissement local et la récidive, en raison des difficultés de l'exérèse, comme par exemple la tumeur desmoïde ou fibromatose agressive.
Tumeurs à malignité locale
Tumeurs dont les caractères histologiques et macroscopiques sont malins mais dont l'agressivité est locale. Le pronostic est plus favorable que ne le laisserait supposer le caractère infiltrant de la tumeur (ex : carcinome basocellulaire de la peau).
Nomenclature des tumeurs
L'examen anatomopathologique d'une tumeur a pour objectif d'établir le type, le grade histologique et le stade (c'est-à-dire l'extension) et d'évaluer le pronostic ; ce qui contribue à déterminer le traitement le plus approprié pour le patient.
La nomenclature des tumeurs suit une terminologie précise (tableaux 05.06, 05.07, 05.08, 05.09, 05.10 et 05.11). Un nom de tumeur se compose généralement d'une racine et d'un suffixe, et peut être associé à un adjectif. La racine définit la différenciation (adéno- désigne une tumeur glandulaire, rhabdomyo- une tumeur musculaire striée, leiomyo- une tumeur musculaire lisse).
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Le suffixe ome est utilisé pour nommer les tumeurs bénignes (adénome, rhabdomyome, leiomyome).
Il existe cependant des exceptions (ex : les lymphomes et les mélanomes sont des tumeurs malignes) :
• granulome (réaction inflammatoire riche en macrophages), tuberculome (masse pseudotumorale de la tuberculose pulmonaire), mycétome (masse pseudotumorale formée de filaments mycéliens) et botriomycome (bourgeon hyperplasique) ne sont pas des lésions tumorales ;
• -matose désigne la présence de tumeurs multiples ou diffuses (angiomatose, leiomyomatose, adénomatose) ;
• carcinome désigne une tumeur maligne épithéliale (ex : adénocarcinome) ;
• sarcome désigne une tumeur maligne conjonctive (ex : rhabdomyosarcome) ;
• blastome désigne une tumeur embryonnaire (ex : néphroblastome ou neuroblastome).
Stade et grade des tumeurs
La classification des tumeurs en fonction de l'organe d'origine et de leur type histologique fournit des informations importantes pour évaluer leur pronostic. Toutefois, d'autres paramètres permettent de préciser le potentiel évolutif. Il s'agit du degré de différenciation (grade) et du degré d'extension (stade) de la tumeur, ainsi que dans certains cas de marqueurs moléculaires.
Grade
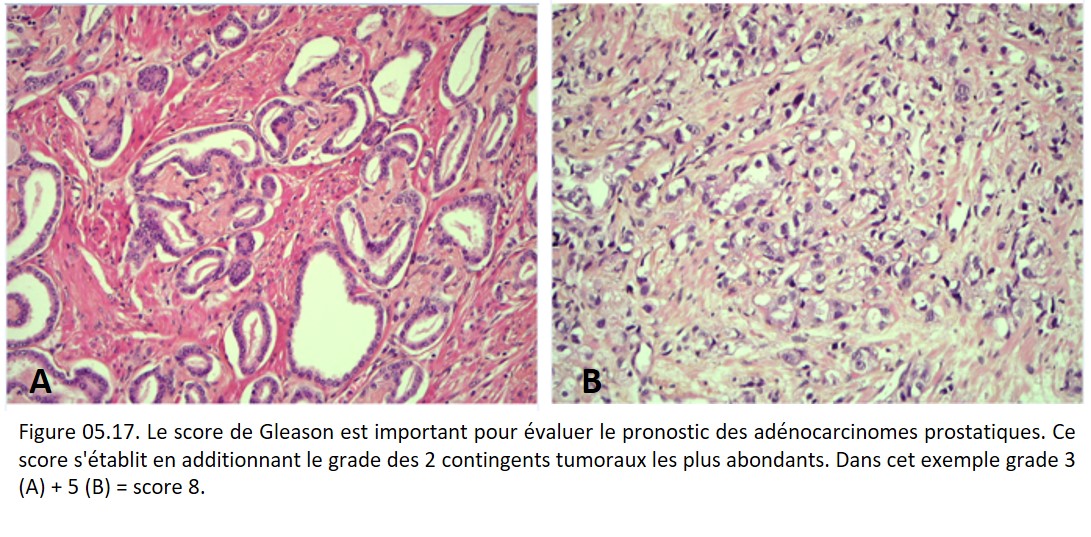
Le grade d'un cancer se fonde sur des critères histologiques tels le degré de différenciation tumorale, l'activité mitotique, le degré d'atypies cyto-nucléaires et/ou l'extension de la nécrose. Il est défini différemment pour chaque type de tumeur (figure 05.17).
{kind=link}
Exemple : le score de Scarff-Bloom-Richardson des adénocarcinomes mammaires (tableau 05.12) prend en compte trois variables : la différenciation glandulaire, les atypies cyto-nucléaires et le nombre de mitoses (comptées sur 10 champs au fort grossissement).
{kind=link}
Stade
La notion de stade et la classification TNM sont traités dans le chapitre 6.
Méthodes diagnostiques des tumeurs
L'étude anatomopathologique a pour but de préciser :
• le type histologique de la tumeur ;
• son agressivité potentielle ;
• son pronostic ;
• sa capacité à répondre à des traitements de plus en plus spécifiques.
Diagnostic morphologique
Après un examen macroscopique attentif permettant de prélever les zones d'intérêt de la prolifération tumorale, la première étape de l'analyse histologique d'une tumeur est l'étape diagnostique. Le diagnostic cyto- ou histologique nécessite de disposer d'échantillons de bonne qualité, relativement abondants et représentatifs de la tumeur et n'ayant pas subi d'altérations pendant leur prélèvement ou leur transport. Les différents modes de prélèvements sont détaillés dans le chapitre 1.
La macroscopie est également cruciale pour déterminer la qualité de l'exérèse d'une tumeur et la présence éventuelle de dissémination dans les tissus non tumoraux (notamment les ganglions lymphatiques).
Examen des coupes histologiques
Les différents prélèvements tumoraux sont examinés dans un premier temps par une coloration standard HE (hémateine/éosine), constituant la base du diagnostic anatomopathologique (typage histologique, grade, stade, limites).
De nombreuses techniques complémentaires, morphologiques ou non, peuvent être utilisées pour confirmer ou préciser le diagnostic. D'autres colorations permettant la mise en évidence des particularités des cellules tumorales (ex. : mucosécrétion avec le bleu Alcian) ou du stroma (ex. : trame réticulinique avec le Gordon-Sweet) sont souvent utiles au diagnostic.
Immunohistochimie
L'immunohistochimie avec des anticorps mono-ou polyclonaux est fréquemment utilisée en pathologie tumorale. L'utilisation de combinaisons d'anticorps dont le choix est orienté par l'étude histologique permet de préciser dans la plupart des cas le type histologique de la tumeur, l'origine primitive des métastases et également la sensibilité à un traitement.
Des anticorps permettent de déterminer la nature des filaments intermédiaires du cytosquelette des cellules. Ces filaments ont une répartition spécifique au sein des grands types de cellules : filaments de cytokératine dans les cellules épithéliales, filaments de desmine dans les cellules musculaires, neurofilaments dans les cellules nerveuses. Ainsi un carcinome est habituellement cytokératine positif (cocktail AE1/AE3) alors qu'un lymphome ne l'est pas.
Les marqueurs de surface sont aussi spécifiques de types cellulaires : antigène CD20 ou CD79a (lymphocyte B), antigène épithélial de membrane (cellules épithéliales), Neural Cell Adhesion Molecule (NCAM) (cellules nerveuses et neuroendocrines), etc.
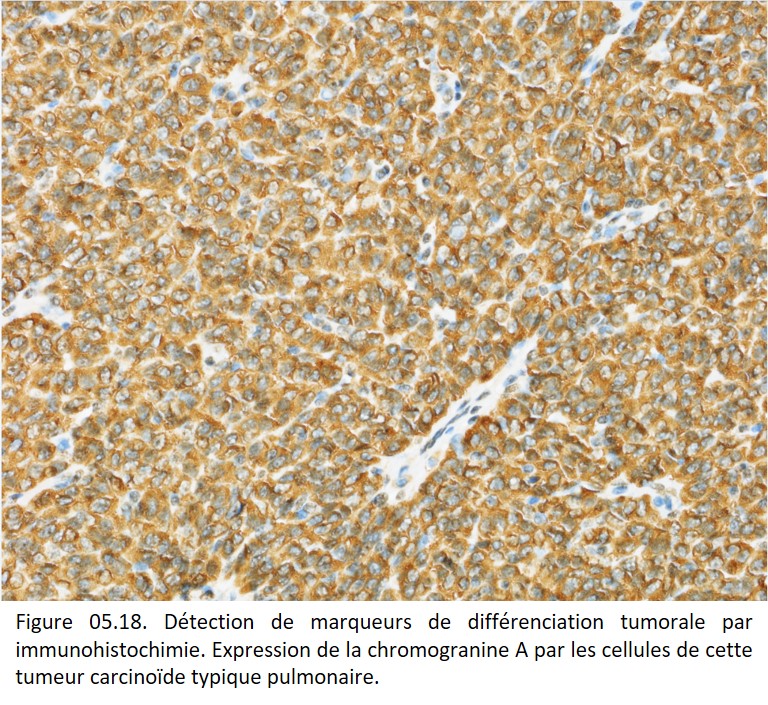
Des marqueurs cytoplasmiques correspondant à des produits de sécrétion ou des molécules fonctionnelles sont aussi exploités : mucines (adénocarcinomes), chromogranine A et synaptophysine (cellules neuroendocrines), HMB45 et Melan A (mélanocytes), thyroglobuline (thyroïde) (figure 05.18).
{kind=link}
Les marqueurs pronostiques seront abordés dans le chapitre 9.
Des anticorps dirigés contre des molécules ayant une valeur pronostique ou thérapeutique sont de plus en plus utilisés. Ainsi, la quantification des récepteurs hormonaux dans les noyaux des cellules tumorales de l'adénocarcinome du sein renseigne sur les effets potentiels d'un traitement anti-hormonal (figure 05.19). La surexpression des protéines ALK, ROS1 ou BRAFV600E donne lieu à un traitement par inhibiteurs spécifiques dans les carcinomes bronchiques non à petites cellules (figure 05.19).
{kind=link}
Pathologie moléculaire
Les techniques de pathologie moléculaire sont utilisées pour mettre en évidence des altérations moléculaires survenues dans les cellules tumorales. Elles peuvent être réalisées sur coupe histologique (ex : hybridation in situ) ou après extraction de l'un des constituants moléculaires du tissu. Les analyses de pathologie moléculaire peuvent être effectuées sur des prélèvements fixés au formol tamponné et inclus en paraffine. L'évaluation histologique préalable de la nature du tissu analysé et de sa richesse en cellules tumorales est indispensable.
Les techniques de pathologie moléculaire ont une valeur diagnostique et pronostique dans certaines tumeurs malignes, et peuvent également aider à prévoir la réponse à une thérapie ciblée (théranostique), à dépister la maladie résiduelle après traitement ou à diagnostiquer une prédisposition héréditaire à développer un cancer.
Les altérations génétiques apparaissent successivement au cours de la croissance d'une tumeur. Certaines de ces anomalies sont récurrentes, c'est-à-dire que le même type d'anomalie survient avec une fréquence élevée dans un type de tumeur donné.
Réarrangements chromosomiques
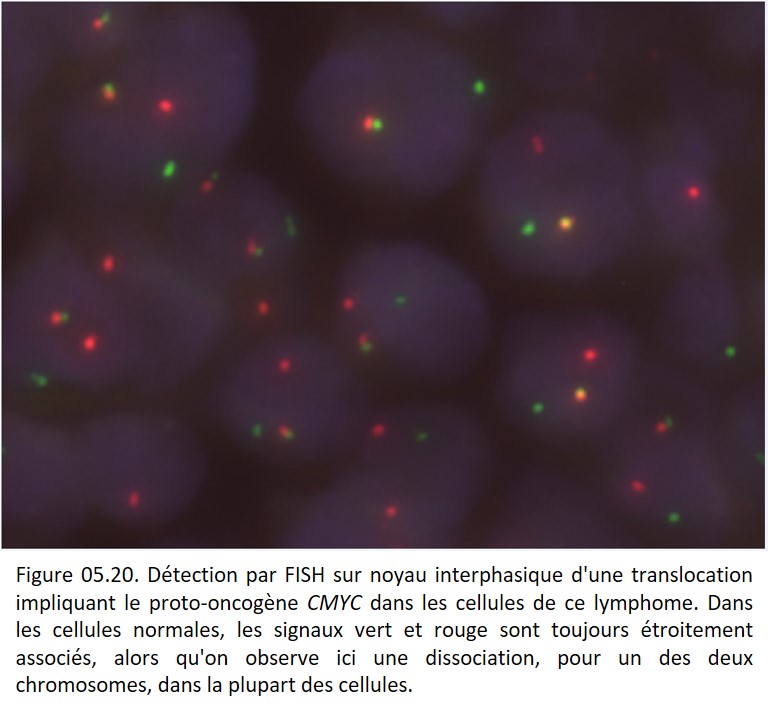
Ces anomalies sont assez fréquentes dans les lymphomes et les sarcomes. Ils peuvent aider au typage des lymphomes (ex : t(14 ;18) des lymphomes folliculaires, t(8 ;14) des lymphomes de Burkitt, t(2 ;5) des lymphomes anaplasiques), des sarcomes (ex. : t(X ;18) des synovialosarcomes) ou des tumeurs pédiatriques (figure 05.20).
{kind=link}
Autres altérations chromosomiques
Il peut s'agir d'anomalies de nombre (ex. : hyperploïdie, aneuploïdie), ou de structure (ex. : l'isochromosome 17q dans les médulloblastomes). Dans les tumeurs à un stade avancé, ces altérations peuvent être très complexes, et différentes d'une cellule à l'autre (sous-clones).
Amplifications géniques
Elles peuvent avoir une valeur pronostique (ex. : mauvais pronostic des neuroblastomes ayant une amplification de CMYC). Elles sont également parfois utiles pour prédire la sensibilité à une thérapie ciblée (ex. : HER2 dans des carcinomes mammaires ou gastriques).
L'instabilité génétique
Dans les adénocarcinomes colorectaux, l'instabilité génétique a été opposée à l'instabilité chromosomique. Elle est liée à un défaut de réparation de l'ADN, qui peut être d'origine héréditaire (syndrome de Lynch) ou acquis (formes sporadiques).
Mutations d'un gène
Elles peuvent être assez spécifiques d'une tumeur (ex : gène KIT pour les tumeurs stromales digestives).
Clonalité d'une tumeur
Elle peut être établie par la mise en évidence du réarrangement clonal des gènes codant pour les immunoglobulines ou pour le récepteur de cellules T (pour les lymphomes) ou par la mise en évidence d'une inactivation clonale du chromosome X (chez les femmes).
Stratégie diagnostique
L'objectif de la prise en charge médicale d'un patient cancéreux est de le traiter le mieux possible, et au moindre coût. Dans la grande majorité des cas, un diagnostic anatomopathologique, avec au minimum un typage de la tumeur, est nécessaire avant le traitement. Toutefois, ceci nécessite le plus souvent un geste invasif qu'il faut mettre en balance avec les risques et l'intérêt pour le patient. Ainsi, chez un patient cirrhotique présentant de volumineux nodules hépatiques et une élévation importante du taux sérique d'alphafœtoprotéine, le diagnostic de carcinome hépatocellulaire est pratiquement certain, et le désagrément et le risque d'une biopsie de confirmation histologique ne sont pas compensés par le bénéfice escompté pour le patient.
Types de prélèvements
Le choix du type de prélèvement est discuté en fonction notamment du patient (état général, antécédents, urgence, souhaits), des hypothèses diagnostiques et des possibilités thérapeutiques.
Prélèvements cytologiques
Par exemple : frottis de lésions cutanéomuqueuses, aspirations bronchiques, cytoponction d'organes profonds. Ils sont les moins invasifs.
Bien que les informations qu'ils apportent soient souvent incomplètes, elles peuvent être rapidement obtenues, et sont souvent utiles pour déterminer le geste le plus approprié pour la suite de la démarche diagnostique (biopsie, chirurgie, chimiothérapie première). Elles sont parfois suffisantes pour décider de la thérapeutique.
Biopsies par voies endoscopiques
Elles sont assez peu invasives et permettent souvent un diagnostic (ex. : typage d'une tumeur bronchique, gastrique ou colique). Le résultat est fiable et l'ensemble du traitement peut être fondé dessus (ex. : lymphome gastrique, carcinome bronchique à petites cellules, adénocarcinome colique).
Biopsies d'organes profonds
Elles constituent une alternative pour les organes pleins (ex. : nodule hépatique ou pancréatique).
Prélèvements chirurgicaux
Ils sont les plus invasifs. Il faut toutefois encore distinguer les prélèvements à visée uniquement diagnostique qui peuvent être de petite taille (ex. : biopsie ganglionnaire ou médiastinale) et les résections à but thérapeutique. Ce sont ces derniers prélèvements qui fourniront le plus d'informations (ex. : le stade d'un adénocarcinome colique dans une iléo-colectomie).
Collaboration au sein de l'équipe médicale
Le médecin qui prescrit ou effectue un prélèvement en vue d'un examen anatomopathologique doit toujours garder à l'esprit que celui-ci doit permettre de faire un diagnostic. Ce prélèvement doit donc être :
• aussi représentatif que possible de la tumeur ;
• acheminé dans les conditions appropriées, c'est-à-dire soit dans du formol tamponné si ce médecin est certain qu'un fragment tumoral frais ne sera pas nécessaire, soit à l'état frais (transport en moins de 30 min et en atmosphère humide) ;
• associé à des informations cliniques qui permettront au pathologiste de le redistribuer pour des techniques appropriées (ex. : biologie moléculaire, caryotype).
En cas de doute, il est souvent utile de discuter avec le pathologiste concerné avant d'effectuer le prélèvement.
Après avoir fait une synthèse des lésions macro- et microscopiques et, le cas échéant, des altérations moléculaires, le pathologiste confronte ses conclusions avec les données cliniques, radiologiques et biologiques afin d'établir un diagnostic définitif.
Définition, composition et classification des tumeurs
• Une tumeur, ou néoplasme, est une masse tissulaire résultant d'une prolifération cellulaire excessive, reproduisant plus ou moins fidèlement un tissu normal ou embryonnaire, et ayant tendance à persister ou croître spontanément. Une tumeur peut être plus ou moins bien différenciée, selon qu'elle ressemble ou non au tissu d'origine.
• Tout cancer est constitué de cellules tumorales (cellules proliférantes anormales) et du stroma qui correspond à l'ensemble des cellules non tumorales présentes en son sein. Les cellules du stroma ne contiennent pas les anomalies génétiques des cellules tumorales.
• Les tumeurs sont classées en fonction de l'organe où elles se développent initialement (= « primitif ») et du type histologique, c'est-à-dire du tissu qu'elles tendent à reproduire. L'examen anatomopathologique doit également évaluer le potentiel évolutif d'une tumeur et notamment, pour les tumeurs malignes, le stade et le grade.
• La classification et la caractérisation des tumeurs nécessitent, dans la majorité des cas, l'examen d'un prélèvement tissulaire biopsique ou chirurgical. Le diagnostic est alors réalisé sur des coupes histologiques colorées à l'HES, complétées si besoin par des techniques complémentaires histologiques (colorations « spéciales », immunohistochimie, hybridation in situ), ou moléculaires réalisées sur l'ADN génomique ou complémentaire.
• La stratégie diagnostique, c'est-à-dire le choix du prélèvement et des techniques les plus appropriées à la situation du patient nécessitent de bien connaître les avantages et les limites de chaque méthode. Une interaction directe entre les médecins ou chirurgiens ayant en charge le patient et le médecin pathologiste réalisant le diagnostic est nécessaire pour le choix de la stratégie diagnostique et du projet thérapeutique.
• La très grande majorité des cancers résulte des effets cumulés d'altérations survenues successivement sur l'ADN des cellules tumorales. Ces altérations sont responsables de la prolifération et l'accumulation de cellules d'origine monoclonale ou oligoclonale, puis du développement de sous-clones.
• Les trois principales familles de gènes impliqués dans la cancérogenèse sont les oncogènes, les gènes suppresseurs de tumeur et les gènes responsables du maintien de l'intégrité du matériel génétique.
• Les altérations responsables de la dérégulation de l'expression ou de la fonction de ces trois grandes familles de gènes peuvent être des anomalies chromosomiques, génétiques ou épigénétiques.
• Les oncogènes et gènes suppresseurs de tumeur sont le plus souvent impliqués dans la régulation des grandes fonctions cellulaires, et notamment le cycle cellulaire, l'apoptose et la signalisation intracellulaire.
• Les facteurs de risque de développement des cancers sont héréditaires (monogéniques ou familiaux) ou environnementaux (virus, radiation, toxique).
• Les interactions entre la tumeur et son hôte (le patient), en particulier l'angiogenèse et la réponse immune, constituent des modes de régulation majeurs de la croissance tumorale.